 |
| Regreso a las bases |
J. García Dorado*, P. Alonso Fraile**
*Dermatólogo. Clínica Dermatológica. Salamanca. **Medicina Familiar y Comunitaria. Clínica Dermatológica. Salamanca
| Resumen
El sistema tegumentario es el órgano más grande del cuerpo y forma una barrera física entre el ambiente externo y el medio interno, sirviendo para protegerlo y mantenerlo en condiciones ideales. El sistema incluye: epidermis, dermis, hipodermis, glándulas asociadas, cabello y uñas. Además de su función de barrera, este sistema realiza muchas funciones complejas como: regulación de la temperatura corporal, mantenimiento de los fluidos celulares, síntesis de vitamina D y detección de estímulos. Los diversos componentes de este sistema trabajan en conjunto para llevar a cabo estas funciones; por ejemplo, la regulación de la temperatura corporal ocurre a través de termorreceptores que conducen al ajuste del flujo sanguíneo periférico, el grado de transpiración y el vello corporal. |
| Abstract
The integumentary system is the largest organ of the body forming a physical barrier between the external and the internal environments, in order to protect and maintain the latter. |
Palabras clave: Epidermis; Dermis; Queratinocito; Melanocito; Alopecia areata; Trastornos del cabello; Tricotilomanía; Onicopatía; Uña.
Key words: Epidermis; Dermis; Keratinocyte; Melanocyte; Alopecia areata; Hair disorders; Trichotillomania; Onychopathy; Nail.
Pediatr Integral 2021; XXIV (3): 156.e1 – 156.e13
Anatomía y fisiología de la piel
La piel conforma la cubierta externa del ser humano, sin soluciones de continuidad, ya que en los orificios naturales, se transforma progresivamente en mucosa. Representa uno de los órganos más importantes del mismo. De un lado por su tamaño, al ser el más grande, ya que cubre una superficie de alrededor de 2 m2 y un peso de 4-5 kg (aproximadamente el 6% del peso corporal total); y por otro, por sus variadas funciones(1) que le permiten, al mismo tiempo, separar al organismo del medio ambiente externo y facilitar su comunicación con él. Aunque la piel del neonato pueda parecer “madura”, atendiendo a su aspecto anatómico, muchas funciones fisiológicas de la piel experimentarán una maduración posnatal(2). Estas competencias se desarrollan y maduran durante toda la gestación y el periodo neonatal; de tal forma, que la piel de un recién nacido pretérmino no es capaz de mantener la homeostasis, ni regular la temperatura y es mucho más susceptible a la infección(3). Estudios recientes han demostrado cómo este desarrollo se mantiene a lo largo del periodo neonatal, siendo en esta etapa la piel muy diferente a la del adulto: escaso desarrollo del estrato córneo, uniones intercelulares epidérmicas más débiles, menor producción de sebo, disminución de la cohesión entre dermis y epidermis, mayor pérdida transepidérmica de agua, la conductancia es mayor, el pH de la piel inferior, piel mucho más fina y con una actividad caseinolítica mucho más alta que en el adulto(4).
La función primordial de la piel es construir una capa córnea eficaz, protectora, semipermeable, que haga posible nuestra supervivencia en el medio en el que nos desenvolvemos. Esta barrera impide tanto la pérdida de fluidos corporales, como la entrada al organismo de elementos nocivos (microorganismos, radiación UV, tóxicos…). Para cumplimentar eficazmente estas funciones protectoras y reguladoras, la piel sintetizará, a lo largo de un complejo proceso de diferenciación, numerosas proteínas y lípidos, incluidos en la composición de la capa córnea.
De forma inseparable, hay que considerar los anexos cutáneos o partes adjuntas a la piel. Además de los pelos y uñas (anexos queratinizados), externos y evidentes, en la piel, se encuentran otro tipo de anexos producidos por invaginaciones de la epidermis: folículos pilosos, glándulas sebáceas y sudoríparas.
Composición de la piel
Constituida por tres capas muy diferentes entre sí en anatomía y función, pero con complejas interrelaciones: epidermis, dermis e hipodermis (Fig. 1).
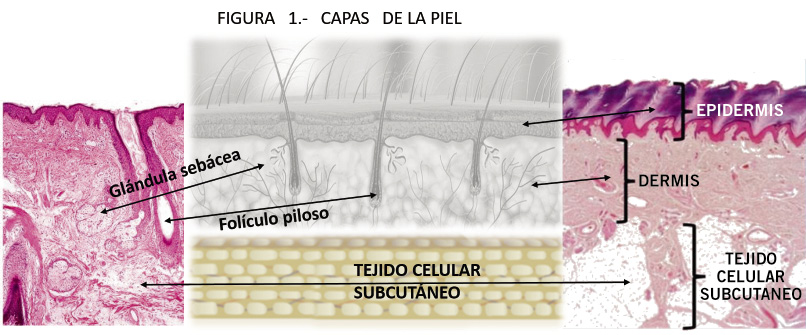
Figura 1. Capas de la piel.
Con origen embriológico totalmente distinto: la epidermis es derivada del tejido ectodérmico, mientras que la dermis y la hipodermis lo hacen del mesodermo, al igual que las células de Langerhans. Por su parte, la cresta neural es responsable de las terminaciones nerviosas sensoriales y de los melanocitos que emigran hacia la epidermis.
Tiene un grosor desde 0,03 (párpado) a 0,12 mm en la piel fina y puede alcanzar 1,4 mm en la gruesa (palmas). El grosor de cada capa varía en función del área anatómica del que hablemos. De esta forma, la epidermis palmar y plantar es la más gruesa al incorporar una capa “extra” denominada estrato lucido.
Las células madre están presentes en todos los niveles de la piel y son pieza clave para entender la fisiología de este territorio(5). Las características inherentes de estas células de auto renovarse y poder dar origen a diferentes tipos celulares, junto con el desarrollo asombroso de la bioingeniería, plantean un horizonte de posibilidades apasionante. En ese sentido, las células madre de la piel tienen un atractivo especial, por su número relativamente elevado, proporcional a la superficie corporal, y su accesibilidad. Se pueden localizar: en la capa basal de la epidermis inter folicular, donde se diferencian hacia queratinocitos; en la parte inferior de la porción permanente del folículo piloso se detectan aquellas responsables de la regeneración del mismo; en la papila dérmica encontramos células con diferenciación hacia el linaje neuronal y mesodérmico; y otras se ubican en glándulas sebáceas, infundíbulo y glándulas sudoríparas.
Epidermis
Es un epitelio plano poliestratificado y queratinizado que cubre la totalidad de la superficie corporal. Es la capa de la piel con mayor número de células y con una dinámica de recambio extraordinariamente grande. Presenta un espesor variable, con un valor medio de 0,1 mm, pudiendo alcanzar en zonas, como las plantas de los pies y las palmas de las manos, espesores de hasta 1 o 2 mm.
Estructura
Predominantemente compuesta por queratinocitos en fases progresivas de diferenciación, que conforme se dividen, se mueven de la capa más profunda a la más superficial, modelando cuatro estratos diferentes que desde el interior hacia el exterior serían (Fig. 2):

Figura 2. Capas de la epidermis.
• Capa basal o estrato germinativo (Fig. 3).
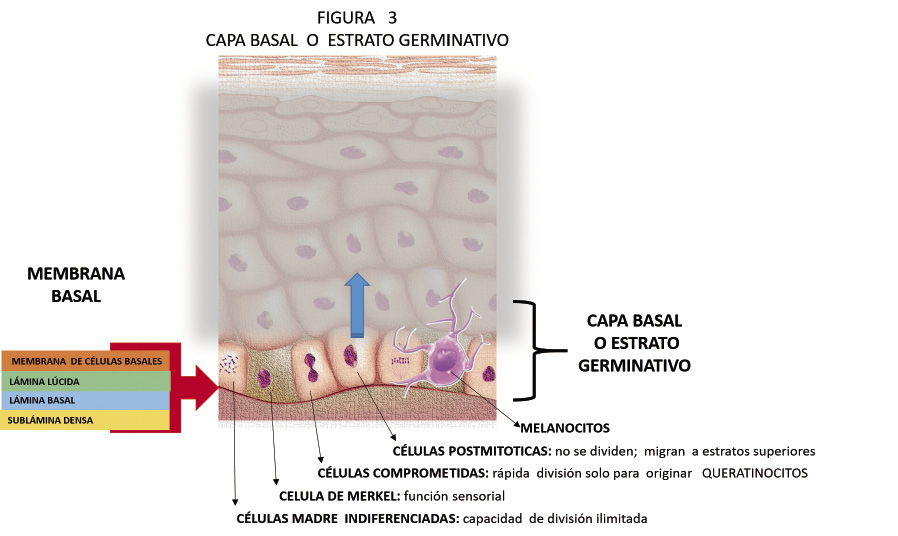
Figura 3. Capa basal o estrato germinativo.
La más profunda. Se encuentra separada de la dermis por la membrana o lámina basal, y anclada a ella por hemidesmosomas. La membrana basal se identifica con facilidad como una membrana homogénea, eosinofílica, PAS+, con una estructura compleja formada por 4 espacios: membrana de las células basales, lámina lúcida, lámina basal y zona fibrosa o sublámina densa. A nivel de las células basales, se pueden observar los hemidesmosomas que unen la epidermis a la lámina densa a través de los filamentos de anclaje, constituidos principalmente por las proteínas: laminina 5 y BP180. La lámina densa está compuesta predominantemente por colágeno tipo IV y está unida a la dermis subyacente por medio de las fibras de anclaje constituidas por colágeno tipo VII.
Sobre la membrana basal, la capa basal queda configurada por una monocapa de células cilíndricas o cúbicas, con núcleo grande. Con tres tipos celulares: células madre indiferenciadas, con capacidad de dividirse de forma ilimitada; células comprometidas, con rápida división, pero solo para originar queratinocitos y células posmitóticas, en estado de diferenciación terminal, que ya no se dividen, sino que migran al estrato superior. Las queratinas K5 y K14 son las principales proteínas del citoesqueleto de los queratinocitos proliferativos de la capa basal. Son sustituidas por las K1 y K10, cuando el queratinocito comienza su proceso de queratinización y se desliga de la membrana basal. La queratina K2 se encuentra en la capa granulosa. También se localizan en esta capa los melanocitos.
• Estrato espinoso o de Malphigio. Contiene 8-10 capas de células “espinosas” poliédricas e irregulares, que se mantienen unidas entre sí por proyecciones en forma de espinas del citoplasma y con puentes intercelulares (desmosomas) por los que contactan con las células vecinas, y constituidos por varias proteínas que se agrupan en: filamentos de queratina, plaquinas y desmogleinas. Son ricas en ADN, necesario para la síntesis proteica que concluirá con la producción de queratina. Aparecen queratinas K1-K10 ocupando el citoplasma. En esta capa también se localizan células dendríticas.
• Estrato granuloso. Aquí se inicia el proceso de queratinización. Constituido por 3-5 capas de células romboidales, más aplanadas, toscamente granulares, en forma de diamante, que contienen gránulos de queratohialina (íntimamente ligados al citoesqueleto celular y necesarios para la producción de queratina K2 y K11) y gránulos lamelares. Estos últimos se sitúan cerca de la membrana plasmática, fuera de la cual vierten su secreción por exocitosis hacia los espacios extracelulares entre el estrato granuloso y córneo. Contienen fosfolípidos, ceramidas y glicolípidos que al depositarse en la superficie de la piel, actúan como un “pegamento” que mantiene las células íntimamente unidas entre sí y forman la probarrera lipídica, que contribuye a la impermeabilización de la piel. A medida que ascendemos en las capas de este estrato, las células van perdiendo los orgánulos citoplasmáticos y quedando solo los desmosomas.
• Estrato lúcido. Selectivamente presentes en palmas y plantas. Con 2-3 capas de queratinocitos diáfanos, muy aplanados que carecen de núcleo y con un citoplasma lleno de una sustancia gelatinosa, la eleidina, precursora de la queratina. Es muy rica en lipoproteínas y su función fundamental es impedir la entrada o salida del agua. Los filamentos intermedios de queratina están íntimamente agregados y se orientan paralelos a la superficie.
• Estrato córneo. El más superficial y ancho de la epidermis, y el que más varía en grosor. Es el resultado de la diferenciación terminal de los queratinocitos epidérmicos. Una capa constantemente renovada y muy protectora. Constituye la primea línea defensiva de la piel y juega un papel clave en la función barrera frente a las agresiones que llegan del medio externo. Está formada por 20-30 capas de células escamosas, muertas, aplanadas y anucleadas denominadas corneocitos, situadas unas sobre otras en forma de tejas y que continuamente se desprenden. Diferenciaríamos dos subcapas dentro del estrato córneo. Una capa córnea compacta, densa, cohesiva, la más profunda; y una capa más superficial, más laxa, menos cohesiva, que a medida que se hace más externa va perdiendo la adhesividad de sus células, se deteriora la adhesión intercorneocitaria y, por último, descaman.
Sin embargo, a pesar de esa aparente inactividad celular, persisten en ella muchos procesos químicos, pre programados en las capas vivas, que desempeñan un papel esencial en la descamación ordenada de los corneocitos superficiales y en la regulación de la permeabilidad del estrato córneo. Estos corneocitos contienen filamentos de queratina inmersos en una matriz de filagrina, de tal modo que la membrana plasmática del queratinocito se va sustituyendo por una envoltura cornificada lipídica. El citoplasma de estas células ha sido sustituido por una proteína hidrófoba denominada queratina. Las uniones entre las células a este nivel aparecen reforzadas, lo que explica la elevada resistencia a la erosión. Las células más superficiales pierden la organización de los desmosomas y su adhesividad, desprendiéndose.
En esta capa, los queratinocitos segregan defensinas, moléculas efectoras de la inmunidad innata con un amplio espectro antimicrobiano e importantes efectos inmunomoduladores, constituyendo parte de nuestro primer sistema defensivo inmune.
Células de la epidermis
Queratinocitos. El grupo celular predominante que constituye el 80% de las células epidérmicas. Se originan en la capa basal, donde son altamente proliferativos y según maduran y ascienden en la epidermis, pierden progresivamente ese potencial replicativo y experimentan una destrucción programada. Es en esta última fase, cuando se tornan queratinocitos anucleados (corneocitos) y contienen solamente filamentos de queratina embebidos en una matriz de filagrina. La descamación es el resultado final. La globalidad de este proceso se denomina queratinización. La renovación de la totalidad de la capa epidérmica se acompaña de transformaciones radicales del queratinocito y se completa en un periodo aproximado de 30 días, desde que se produce la división celular hasta que la célula cae desprendida de la superficie de la piel.
A lo largo de todo este ciclo y según la fase de diferenciación, la célula sintetiza:
• Varios tipos de queratinas, que son una familia de proteínas estructurales principales de los queratinocitos. Insolubles en agua y con una gran resistencia frente a cambios en el pH y a elevadas temperaturas. También presentan una fuerte resistencia a la degradación enzimática. El aminoácido principal de la queratina es la cisteína. Químicamente, tienen unos puentes di-sulfuro (azufre) que dan fuerza y rigidez a la estructura, dependiendo de la cantidad de puentes será más dura o menos. Globalmente, se subdividen en dos grupos, las queratinas duras o ? (alfa) que forman parte del pelo y uñas; y las blandas o ? (beta) que son los elementos esenciales de la capa córnea.
• También sintetiza diversos lípidos que permiten controlar la permeabilidad de la epidermis. Esta importante síntesis proteica, la formación de numerosas uniones intercelulares y el constante reciclaje de las células, permiten mantener una epidermis suficientemente elástica, cohesiva e impermeable.
La queratinización muestra diferentes etapas:
• La división celular ocurre en el estrato basal o germinativo. Fruto de esa mitosis, una célula cuboidal permanece y la otra asciende hacia la superficie de la piel, y ya comienza a sintetizar tonofilamentos (compuestos de queratina), que se agrupan en haces que constituyen las tonofibrillas.
• La célula llega al estrato espinoso. Y al incorporase a la parte más superior de esta capa, las células comienzan a producir gránulos de queratohialina que contienen proteínas intermedias, filagrina y tricohialina. También producen cuerpos lamelares.
• Desde ahí, son empujadas hacia el estrato granuloso, donde se aplanan y adoptan la forma de diamante. Estas células acumulan gránulos de queratohialina mezclados entre las tonofibrillas.
• Por último, las células avanzan hacia el estrato córneo, donde se aplanan y pierden las organelas y el núcleo. Los gránulos de queratohialina transforman las tonofibrillas en una matriz homogénea de queratina.
• Finalmente, las células cornificadas alcanzan la superficie de la piel: la envoltura cornificada-lipídica sustituye a las membranas plasmáticas de los queratinocitos previos; las células se aplanan, se interconectan entre sí mediante corneosomas y se amontonan como capas, conformando el típico aspecto del estrato córneo. Queda configurada esta barrera más externa a expensas de una red de corneocitos y matriz lipídica extracelular, y son descamadas a través de la rotura de los desmosomas.
En cuanto a la maduración intraútero, a las 21 semanas de gestación, el estrato córneo ya está formado; a las 28 semanas ya tiene 2-3 capas de células y a las 32 ya se detectan, al menos, 15 capas, equivalentes a la piel adulta. A los 6 meses de embarazo, la epidermis ya funciona como una barrera entre el feto y el medio exterior. En nacidos por debajo de las 32 semanas, el estrato córneo es muy fino y no previene suficientemente la pérdida transepidérmica de agua, la absorción de tóxicos externos y la invasión de gérmenes. Pero la exposición al medio ambiente tras el nacimiento en prematuros, acelera y estimula la maduración. Ese estrato córneo madura muy rápido hasta el final de la primera semana de vida, pero alcanzar la maduración total le llevará más de 4 semanas. Si la edad gestacional es menor de 25 semanas, el estrato córneo tardará 8-10 semanas en madurar; mientras que en edades gestacionales de 27-28 semanas, se alcanza la maduración total en unos 10 días(6).
Melanocitos (Fig. 4).
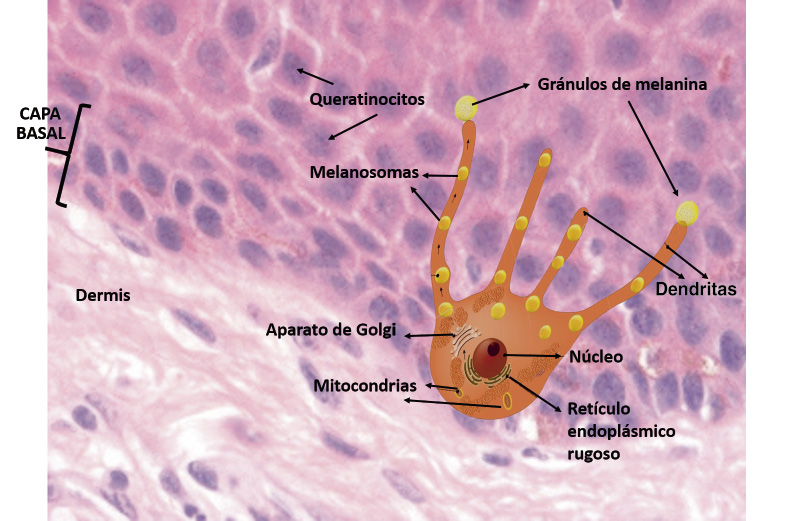
Figura 4. Melanocitos.
Células dendríticas derivadas de la cresta neural, desde donde migran para asentarse entre las células basales de la epidermis y el folículo piloso. Suponen alrededor del 10% de las células epidérmicas y son responsables de la producción de melanina que, a su vez, condiciona la pigmentación de la piel. La luz UVB estimula la formación de melanina, que actúa como un “fotoprotector” natural. Se suelen disponer en la basal epidérmica y contactan con los queratinocitos a través de sus dendritas, existiendo un melanocito por cada 36-40 queratinocitos (unidad melánica epidérmica) o un melanocito por cada 9 células basales. El melanocito posee unas prolongaciones de su citoplasma donde se forma la melanina (melanogénesis). Diferenciamos dos tipos de melanina: la de color marrón parduzco (eumelanina) y la de color rojo amarillento (feomelanina). Su síntesis está regulada por diversas enzimas, entre las que destaca la tirosinasa, que metaboliza el aminoácido tirosina para formar dihidroxifenilalanina (DOPA). La actividad de la enzima está estimulada por la unión de la MSH (melanocite stimulating hormone) a un receptor en la membrana de los melanocitos. Y, a su vez, este receptor MCR (melanocortin receptor) presenta hasta 5 variantes y, dependiendo de la variante presente, se va a determinar la respuesta en la producción de un tipo u otro de melanina (eumelanina o feomelanina), lo que va a determinar la respuesta del individuo a la luz solar.
Una vez formada la melanina, la transfieren a los queratinocitos adyacentes en forma de melanosomas. Los diferentes tonos de la piel son debidos a: diferencias en la actividad de los melanocitos, las características de los melanosomas, así como la capacidad de transferir este pigmento a los queratinocitos, cantidad de la melanina, el tamaño y densidad de los melanosomas transferidos, más que al número de los melanocitos.
Células de Langerhans (Fig. 5).
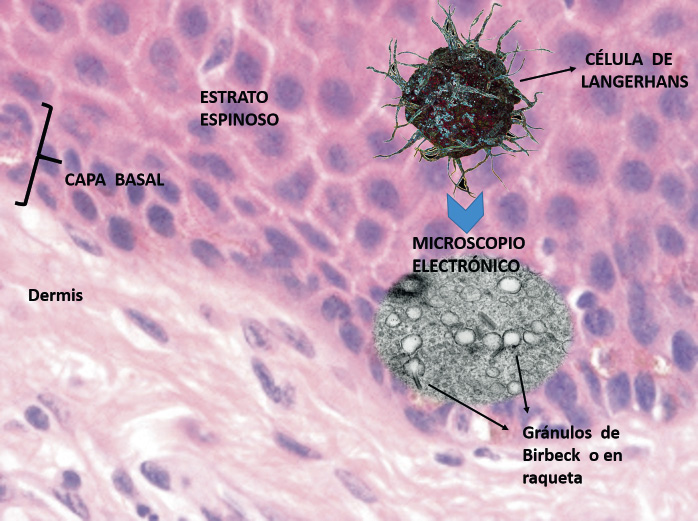
Figura 5. Célula de Langerhans.
Estas células dendríticas conforman la primera línea defensiva de la piel y juegan un papel clave en la presentación de antígenos. Se localizan en el estrato espinoso y representan el 3-8% de la población celular de esa capa. Son de origen mesenquimal, derivadas de las células madre CD34 positivas de la médula ósea, forman parte del sistema mononuclear fagocítico y están involucradas en una gran variedad de respuestas inmunes por medio de la activación de las células T. Las células de Langerhans tienen una distribución muy constante en toda la piel y pueden detectarse por medio de la localización de diversos antígenos como: ATPasa, CD1a, langerina, CD4, S100 y HLA-DR. En el citoplasma, las células contienen un gránulo característico que se observa en microscopía electrónica y que tiene forma de gusano o raqueta, conocido como gránulo de Birbeck. Estas células expresan moléculas MHC I y MHC II, captan antígenos en la piel que se procesan en compartimentos especializados y un fragmento de los cuales se une a complejos de histocompatibilidad mayor. Tras unas horas, las células aumentan de tamaño, abandonan la epidermis, migran a través de la dermis y entran en los vasos linfáticos dérmicos y, de ahí, hacia las áreas paracorticales de los ganglios linfáticos de drenaje, en donde presentan el antígeno a las células T, dando lugar a una respuesta específica y productiva en estas células.
Células de Merkel. Son células epidérmicas modificadas, que provienen de la cresta neural y se localizan en el estrato basal, directamente sobre la membrana basal. Tienen una función sensorial como mecano-receptores, estando situadas en lugares con sensibilidad táctil muy intensa como son: pulpejos, palmas, plantas y mucosa oral o genital. Están unidos a través de desmosomas con los queratinocitos adyacentes y contienen filamentos de queratina intermedia. Su membrana interactúa con las terminaciones nerviosas intraepidérmicas. La queratina 20 es el marcador más eficaz de la célula de Merkel.
Dermis
Es la estructura de soporte de la piel y le proporciona resistencia, elasticidad y capacidad de adaptación a movimientos y cambios de volumen. De origen mesenquimal, constituye la mayor masa de la piel y su grosor máximo es de unos 5 mm. Conexiona con la epidermis a nivel de la membrana basal. Aunque contiene elementos celulares, la composición es principalmente fibro-elástica, conteniendo colágeno y fibras elásticas. Entre el componente fibroso, la matriz extracelular contiene una elevada proporción de glicosaminoglicanos, tales como: ácido hialurónico, proteoglicanos y glicoproteínas. Es un tejido vascularizado que sirve de soporte y alimento a la epidermis. En ella están inmersos, además: vasos sanguíneos, linfáticos, nervios sensitivos, músculos, folículos pilosebáceos y glándulas sudoríparas.
Componentes de la dermis
Células de la dermis (Fig. 6).
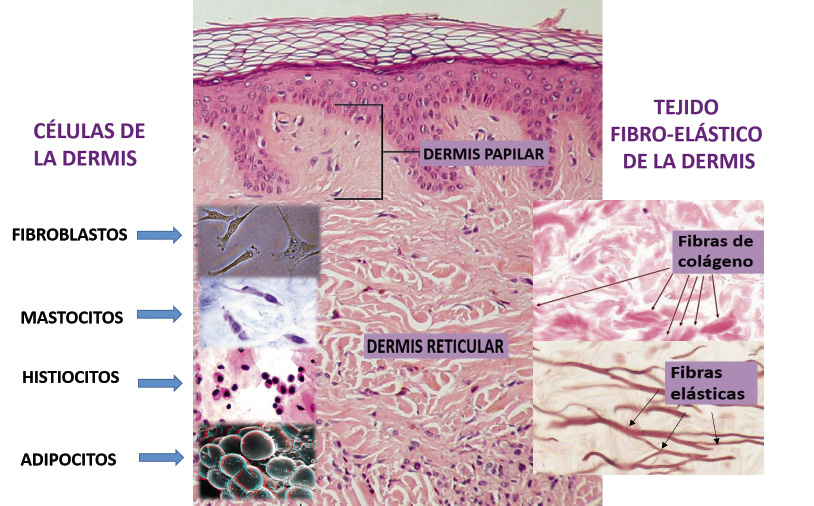
Figura 6. Células de la dermis.
Son escasas e incluyen: fibroblastos, histiocitos, mastocitos o células cebadas, adipocitos, linfocitos, células plasmáticas, eosinófilos y monocitos. Los fibroblastos representan la célula fundamental y se diferencian en fibrocitos, que al enlazarse unos a otros forman un entramado tridimensional. Sintetizan y liberan los precursores del colágeno, elastina y proteoglucanos para construir la matriz extracelular. Los histiocitos son macrófagos tisulares que se implican en el sistema inmune. Los mastocitos son células inflamatorias localizadas en las áreas perivasculares de la dermis que segregan mediadores vasoactivos y proinflamatorios implicados en la respuesta inflamatoria, remodelación del colágeno y curación de las heridas. Los adipocitos dérmicos son diferentes a los localizados en el tejido adiposo subcutáneo. Están implicados en: aislamiento de estructuras dérmicas, depósito de energía, regeneración del folículo piloso y curación de las heridas.
Matriz extracelular o sustancia fundamental. Representa el espacio libre entre los elementos celulares y fibrosos. Está relleno con un fluido gelatinoso, en el que las células se pueden mover libremente. Es una sustancia amorfa compuesta principalmente por proteoglucanos (heteropolisacáridos y proteínas), que debido a su gran capacidad de absorción de agua, forman una materia pegajosa y gelatinosa, que no solo sirve como elemento de unión entre el resto de elementos, tanto celulares como fibrosos, sino que influencia la migración, la cementación y la diferenciación celular.
Tejido fibro-elástico (Fig. 6). El colágeno es el principal componente de la dermis. No es homogéneo en todo el organismo, existiendo 13 tipos en relación con: morfología, composición de aminoácidos y propiedades físicas. La dermis contiene principalmente colágeno tipo I (85-90%), tipo III (8-11%) y tipo V (2-4%). También tienen un papel clave las fibras elásticas. Son esenciales para las propiedades retráctiles de la piel y solo representan un 2-4% de los constituyentes de la dermis. Están compuestas por elastina y microfibrillas de fibrilina. La fibrilina es una glucoproteína esencial para la formación de las fibras elásticas del tejido conectivo. La fibrilina-1 es el mayor componente de las microfibrillas, que constituyen un armazón sobre el cual se deposita la elastina. La especial configuración bioquímica de la elastina le permite, a diferencia del colágeno: desplazarse, elongarse y retraerse. Hay dos subtipos de fibras elásticas, consideradas inmaduras: las fibras de elaunina, dispuestas horizontalmente a nivel de la unión de la dermis reticular y papilar; y las fibras oxitalánicas, dispuestas perpendicularmente en la dermis papilar.
Histológicamente, la combinación del tejido fibroelástico, células dérmicas y matriz extracelular en la dermis, conforma dos áreas diferentes, pero sin clara demarcación entre ellas:
1. La capa papilar o dermis papilar, la más superior, más fina, que contacta con la epidermis y se compone de tejido conectivo laxo. Recibe ese nombre por la presencia de proyecciones hacia el interior de la epidermis. Estas proyecciones se denominan papilas dérmicas y se alternan con los procesos interpapilares de la epidermis. En las papilas se encuentran las asas capilares (sistema circulatorio) que proporcionan los nutrientes a la epidermis avascular. La capa papilar también contiene numerosas terminaciones nerviosas, receptores sensoriales y vasos linfáticos.
2. La dermis reticular, más profunda, más gruesa, menos celular, con tejido conectivo muy denso. Recibe ese nombre por el entramado o retícula de las fibras colágenas que forman gruesos haces entrelazados con haces de fibras elásticas.
Tejido subcutáneo o hipodermis
Está formada por tejido adiposo que forma lobulillos separados por tabiques de tejido conectivo, continuación del conectivo de la dermis reticular, sin un límite definido. Se trata de tejido conectivo laxo y muchas de sus fibras se fijan a las de la dermis, formando puntos de anclaje, fijando así la piel a las estructuras subyacentes (fascia, periostio o pericondrio). Si estos puntos de fijación están poco desarrollados, la piel se desplaza formando plegamientos. Si están muy desarrollados o son muy numerosos, como es el caso de la planta de los pies o del cuero cabelludo, la piel es casi inamovible. El grosor de la hipodermis es muy variable, dependiendo de la localización, el peso corporal, el sexo o la edad. En su espesor, también podemos detectar folículos pilosos, nervios sensitivos y vasos sanguíneos.
Las características de este tejido graso pueden variar dependiendo de la madurez de la piel. Alrededor de la semana 26 de gestación comienza ya la formación del tejido graso subcutáneo y va avanzando paralelamente al embarazo. En los recién nacidos, la grasa subcutánea es rica en ácidos grasos saturados (palmítico y esteárico). Tiene un punto de fusión más elevado que los ácidos insaturados propios de la grasa los adultos. Esto explicaría la mayor facilidad de congelación del tejido graso en los niños que en los adultos y, con ello, determinadas patologías del panículo en ese grupo de edad (necrosis grasa subcutánea del recién nacido, paniculitis por “ingesta de helados”…). Por ello, sería relevante evitar temperaturas extremas en los primeros años de la vida. La grasa parda se localiza en profundidad, en la base del cuello, tras el esternón y área perirrenal, comenzándose a formar a partir de la semana 17-20. Es un tejido adiposo especializado que produce calor mediante la oxidación de ácidos grasos. En prematuros, está incompletamente desarrollada.
Estructura vascular y linfática de la piel
Ubicadas en la dermis. El aporte vascular se dispone en dos plexos (Fig. 7).
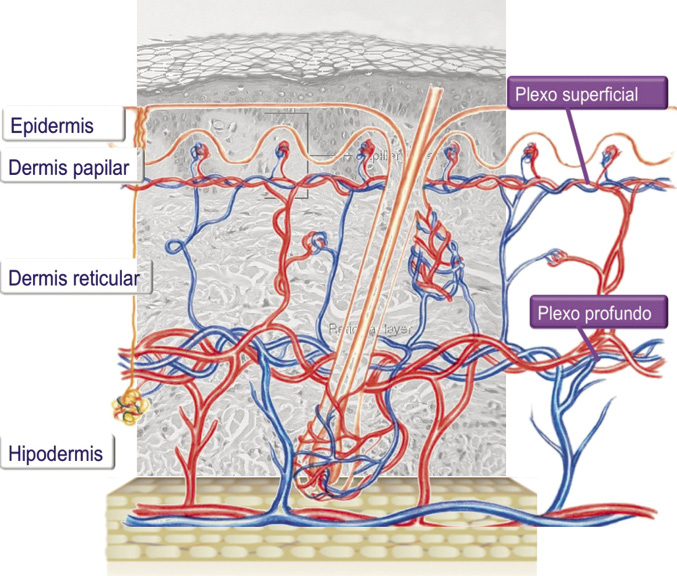
Figura 7. Red vascular de la piel.
El más superficial discurre entre la dermis papilar y reticular. Mientras que el segundo se dispone entre la dermis y la hipodermis. A pesar de esta “simplicidad” anatómica, la fisiología de esta microcirculación y los cambios en el flujo sanguíneo cutáneo son muy complejos fruto de la interrelación de varios factores(7):
• Activación simpática, que produce vasoconstricción a través de liberación de norepinefrina, neuropéptido Y y ATP.
• El sistema colinérgico simpático está implicado en la vasodilatación, mediante la participación de: acetilcolina, péptido vasoactivo intestinal y polipéptido activador de la adenilato ciclasa hipofisaria, también conocido como PACAP.
• Una vasomodulación endotelio-dependiente implica al óxido nítrico, prostaciclinas, factor hiperpolarizante derivado del endotelio y endotelina.
• Respuesta miogénica que juega igualmente un papel definido en esta regulación. El aporte de la epidermis es a través del plexo arteriovenoso superficial (plexo subepidérmico-papilar), vasos clave para la regulación de la temperatura. Se trata de un mecanismo altamente efectivo de regulación de la temperatura a través del tegumento, incrementando el flujo sanguíneo en la piel, transfiriendo con ello el calor desde el organismo hacia el entorno exterior. A su vez, los cambios en el flujo sanguíneo se controlan por el sistema nervioso autónomo: la estimulación simpática conlleva vasoconstricción y, por ello, retención de calor; por su parte, la vasodilatación implica pérdida de calor. Esta vasodilatación es la respuesta al incremento de temperatura corporal, a través de la inhibición de los centros simpáticos del hipotálamo posterior.
En el 5º mes de gestación, ya se reconoce una diferenciación en arteriolas, vénulas y capilares, pero la completa maduración no se produce, sino tras el nacimiento.
Estructura nerviosa de la piel
El sistema nervioso periférico, tanto autónomo como somático, discurre por el espesor de la piel. Existe un sistema eferente, representado por el sistema nervioso autónomo simpático, que es responsable del funcionamiento del sistema vascular y anexial: tono de la vasculatura, estimulación pilomotora de la raíz del cabello y de la sudoración. Y un sistema aferente, el somático sensorial, responsable de procesar información sensorial a través de diferentes receptores sensoriales (Fig. 8).
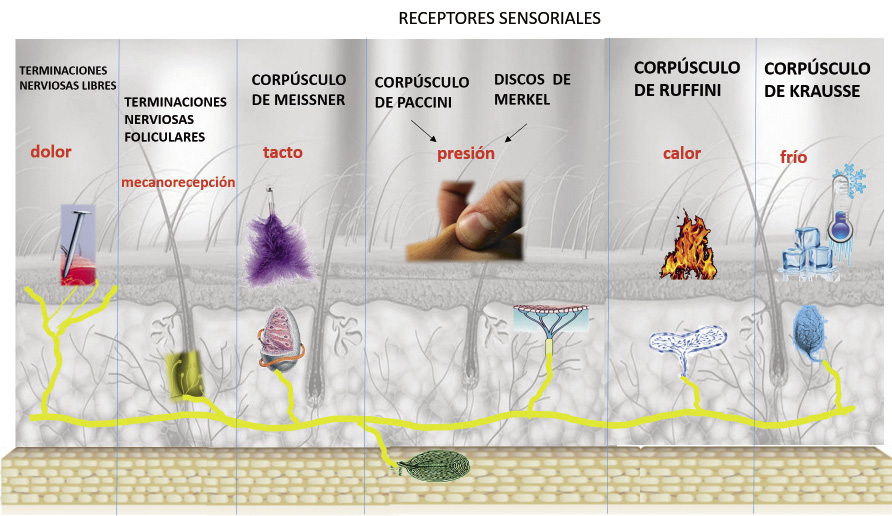
Figura 8. Receptores sensoriales.
Las terminaciones nerviosas son ya visibles a partir del 4º mes. Y ya se puede constatar una respuesta del feto al tacto y al dolor a partir de las 20 semanas. En el RN ya están bien desarrolladas, aunque posteriormente completaran su maduración.
Músculos en la piel
Representado en la piel por el músculo arrector del pelo en forma de fascículos de fibras de músculo liso, sujetos al tejido conectivo de la vaina folicular. Cuando se contrae el músculo, conlleva la erección del vello, no de forma totalmente perpendicular, sino con una amplia angulación y al mismo tiempo comprime la glándula sebácea, desencadenando la secreción de su contenido. Esta “erección capilar” da a la piel el característico aspecto de “piel de gallina”.
Funciones de la piel
Función de barrera epidérmica
Mediante la construcción de la capa córnea, la función de «barrera» vital de la epidermis garantiza el mantenimiento del medio fisiológico interno y protege el organismo contra las agresiones ambientales (calor, frío, radiaciones UV…), la penetración de sustancias potencialmente dañinas y la colonización por bacterias patológicas. Sin menospreciar que garantiza una apariencia saludable y una función adecuada de toda la piel. Ya está presente en el RN a término, aún con diferencias que ya comentaremos. Pero en el RN pretérmino es tan incompleta que la mortalidad, generalmente por causas microbianas, está aumentada. Es directamente proporcional al deterioro de la función misma. Y a mayor prematuridad, paralelamente más facilidad para la absorción de productos químicos.
Esta barrera se configura:
• Por una envoltura celular, con corneocitos que se asocian mediante uniones especializadas (corneodesmosomas), responsables del refuerzo mecánico, que protegen la actividad mitótica de capas subyacentes del daño UV, regula el inicio de la inflamación citoquina-dependiente y mantiene la hidratación.
• Una capa de proteínas insolubles en la superficie interna de la membrana plasmática, que refuerza los mecanismos de barrera. Formada por enlaces cruzados de pequeñas proteínas ricas en prolina y otras más grandes como: cistatina, desmoplaquina y filagrina.
• Una envoltura lipídica, una capa lípido-hidrofóbica, anclada a la superficie más externa de la membrana plasmática, que: regula la permeabilidad, inicia la descamación córnea, tiene actividad peptídica antimicrobiana, elimina toxinas y permite una absorción química selectiva.
Durante la vida fetal, la función barrera es crítica en el sentido de intercambio de agua y gases, y experimenta una drástica modificación tras el nacimiento. Aún hay aspectos sin clarificar, como son: papel exacto de la vérnix caseosa, cómo se implanta la microbiota fisiológica o papel exacto de las acuoporinas, proteínas que actúan como canales transmembrana, pertenecientes a la familia MIP y que forman poros en las membranas biológicas, por los que transportan principalmente agua(8). Al parecer, estos cambios adaptativos, configuración de lo que ya será el estrato córneo, con su función de barrera y de intenso desarrollo, se gestionan desde momentos antes del nacimiento hasta los 4 años de edad, en que adquieren ya valores de adulto(9). En el RN pretérmino, la completa maduración de la función barrera se completa entre las 2 y 4 semanas de la vida posnatal.
La permeabilidad epidérmica es mayor cuanto menor es la edad gestacional. En el RN a término, la permeabilidad es equivalente a la del adulto; por ello, los productos de aplicación tópica pueden alcanzar concentraciones sanguíneas elevadas, debido al aumento de la relación entre la superficie cutánea y el peso corporal.
Aparte de estas diferencias estructurales, otra serie de factores convergen para hacer que la piel del niño en los primeros momentos de su vida más susceptible a la toxicidad percutánea: alta relación superficie corporal/volumen, inmadurez de su capacidad para el metabolismo de las drogas, y su mermado depósito en tejido graso subcutáneo. Por ello, se incrementa el área absortiva y, tras la absorción, el niño carece de sistemas totalmente desarrollados para el transporte de las toxinas y sistemas de desintoxicación desarrollados. Como conclusión directa: parece prudente recomendar en los primeros meses de vida, la aplicación tópica de solo los productos necesarios.
Función de soporte y protección
Asumida en gran parte por la dermis, ya que su complejo diseño la configura como el esqueleto perfecto, que aporta al mismo tiempo: flexibilidad, fuerza y protección de estructuras anatómicas más profundas. Tanto el colágeno como el ácido hialurónico, fortalecen la piel y proporcionan un anclaje sólido de la epidermis vía hemodesmosomas y otros componentes adhesivos de la zona de membrana basal. Igual papel de anclaje epidérmico juegan las fibras oxitalánicas. Por su parte, las fibras elásticas confieren la flexibilidad. El entramado vascular dérmico, facilitando la llegada de nutrientes, es esencial para mantener tanto a la epidermis como sus apéndices. Y la hipodermis que sirve de almacén de energía, además de aislante térmico y de protector mecánico frente a golpes.
Función inmunitaria
Conexionada con la anterior de barrera, e intenta impedir la entrada de patógenos(10). Los péptidos antimicrobianos (AMPs) y los lípidos actúan como una barrera biomolecular que altera las membranas bacterianas. Los AMPs, tales como defensinas y catelicidinas, son producidos por diferentes células, como células dendríticas o macrófagos. Los lípidos, tales como esfingomielina y glucosiceramidas, se almacenan en los cuerpos lamelares del estrato córneo y muestran actividad antimicrobiana. Otro aspecto es el relacionado con la inmunidad celular. Células mieloides y linfoides están presentes en la piel y algunas, como las células de Langerhans, poseen la capacidad de viajar hacia la periferia y activar el resto de sistema inmune.
Función endocrina
La vitamina D3 o colecalciferol se obtiene principalmente de dos fuentes básicas: la dieta (10%) y la producción endógena por conversión fotoquímica a partir de 7-dehidrocolesterol en la epidermis, concretamente por el queratinocito, con la participación de la luz UV del sol. Posteriormente, el colecalciferol es hidroxilado en el hígado y riñón hacia su forma activa de 1,25 dihidroxi vitamina D (calcitriol), que será el responsable de incrementar la absorción del calcio en el intestino.
Función exocrina
Viene dada a través de la secreción del sudor y de las glándulas sebáceas,
Función en la curación de las heridas
Proceso complejo en el que se implican diversos componentes para completar las diferentes fases de: hemostasia, inflamación, proliferación y remodelación(11). En la hemostasia están implicados factores tisulares, presentes en los espacios subendoteliales de la piel, encargados de desencadenar la cascada de la coagulación hasta formar el coagulo de fibrina. En la fase inflamatoria, son los neutrófilos y los monocitos los que acuden a la zona dañada para eliminar patógenos y detritus. La proliferación de queratinocitos y fibroblastos son los responsables de la fase proliferativa, contribuyendo a la formación del tejido de granulación. Y, por último, la participación de los macrófagos, es esencial en la fase de remodelación final: segregan una matriz de metaloproteasas que elimina el exceso de colágeno y mantiene colágeno inmaduro para finalizar la matriz extracelular.
Función de termorregulación
Participada especialmente por el entramado vascular vasoactivo dérmico, visto con anterioridad. También están implicadas unas estructuras especializadas llamadas cuerpos glómicos, constituidos por: células glómicas, vasos y células musculares lisas.
Función sensorial
Llevada a cabo por la existencia de numerosas terminaciones nerviosas, que contienen receptores para: tacto, calor, frío, presión, vibración y dolor.
Anejos de la piel
Aparato ungueal
Consta de estructuras especializadas y un producto córneo: la lámina ungueal (Fig. 9).
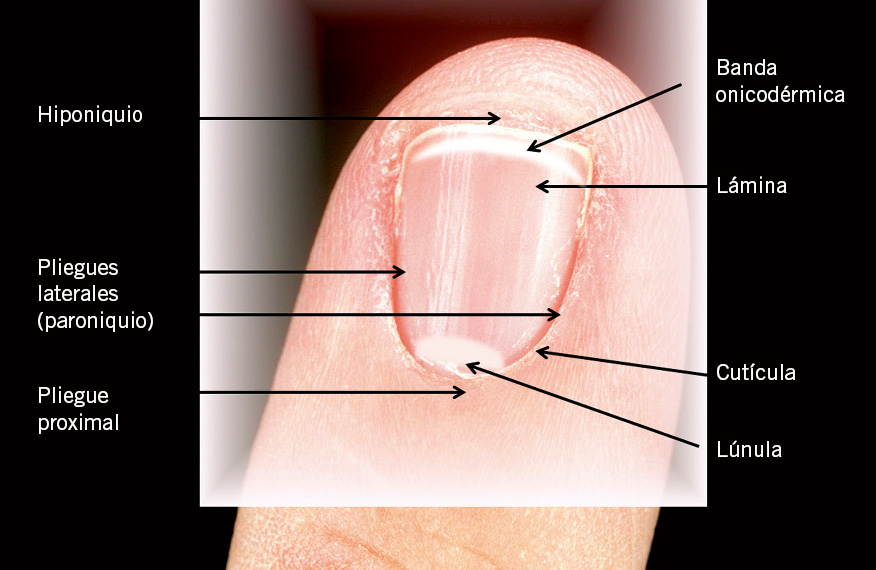
Figura 9. Anatomía de la unidad ungueal.
La lámina ungueal es producida por la matriz ungueal, la cual descansa bajo el pliegue ungueal proximal. Según crece la lámina ungueal, emerge bajo el citado pliegue proximal y progresa distalmente, apoyada sobre el lecho ungueal, con el que establece una íntima adherencia, hasta llegar al pliegue ungueal distal. La superficie de contacto con la matriz ungueal es de 15-25%, mientras que con el lecho ungueal es de 75,85%. Crece durante toda la vida.
La lámina ungueal es una estructura queratinizada, semitransparente y dura, que está formada por tres láminas horizontales: fina capa dorsal, lámina intermedia –más gruesa–, y una cara ventral que contacta con el lecho ungueal. Son células escamosas aplanadas íntimamente unidas entre sí. En la cara dorsal e intermedia existen gran cantidad de fosfolípidos, por lo que aquellos medicamentos con carácter lipofílico, pudieran depositarse en esas capas vía matriz ungueal y provocar discoloración.
Su forma es curva, tanto en el eje longitudinal como en el transversal, lo que le confiere fuerte adherencia a pliegues laterales, proximal e hiponiquio, y que se traduce en una gran resistencia frente a los traumatismos (Tabla I). La superficie externa muestra crestas que varían con la edad.

La composición es muy peculiar, con variaciones en relación con sexo y edad (Tabla II).

El lecho ungueal soporta gran parte de la lámina y abarca desde el pliegue ungueal proximal hasta el hiponiquio. Es un epitelio delgado, con baja tasa de proliferación, con queratinas diferentes a las de piel normal y diferentes a las de la matriz ungueal(12).
Carece de capa granulosa y presenta células paraqueratósicas adheridas fuertemente a la lámina ungueal(13). La dermis subyacente no exhibe estructuras anexiales foliculares ni glándulas sebáceas. Escaso tejido adiposo y una unión muy firme al periostio, a través de potentes fibras de colágeno.
En la parte más distal del lecho ungueal, encontramos una banda transversal de 1-1,5 mm, con un color característico, según el fototipo de piel. Es el denominado istmo o banda onicodérmica, rosada en caucásicos y marrón en afrodescendientes(14). Muestra unos queratinocitos claros nucleados, con una queratinización especial. Y protege de la entrada de agentes externos por debajo de la lámina ungueal.
Proximal y lateralmente, la lámina ungueal está rodeada por los pliegues ungueales (PU). El PU proximal o posterior, representa la continuidad de la epidermis, y la dermis del dorso del dedo. Contiene glándulas sudoríparas, pero no unidades pilosebáceas. Consta de dos capas de epitelio: capa dorsal (que viene a ser la continuación de la piel que recubre el dorso del dedo) y capa ventral, la cual se continua con la matriz ungueal.
Los PU laterales rodean íntimamente a la lámina ungueal y sellan sólidamente los laterales, para protegerlos de la entrada de material extraño. Carece también de estructuras pilosebáceas.
La cutícula tiene como misión proteger la matriz ungueal. En realidad, es una prolongación del estrato córneo de la parte dorsal y ventral del PU proximal.
Formada por tejido cornificado, está adherida íntimamente a la lámina ungueal.
Por su parte, el eponiquio sería la continuidad de la porción ventral del pliegue ungueal proximal. Es una zona sensible, en cuanto a la formación adecuada de la lámina.
El hiponiquio marca el final del lecho ungueal y corresponde al borde libre distal del aparato ungueal. Es la zona de unión de la lámina ungueal con el pliegue ungueal distal. A este nivel, la queratinización es ya similar al de la piel normal, apareciendo ya, de nuevo, capa granulosa.
La matriz ungueal (Fig. 10) es la responsable de la producción de la lámina ungueal, y consiste en un epitelio germinativo, que queratiniza mediante un proceso denominado onicoqueratinización, sin la formación de capa granulosa.
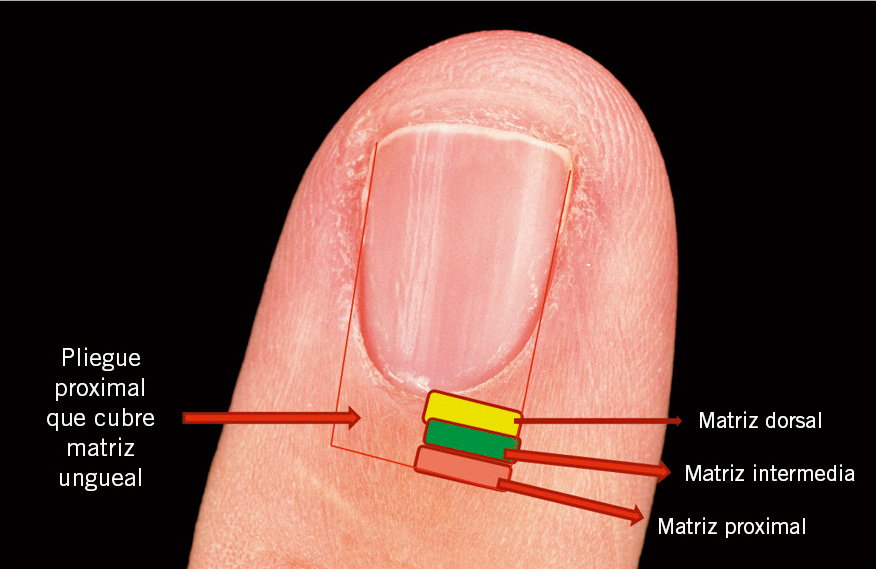
Figura 10. Anatomía de la matriz ungueal.
Este característico y único patrón de queratinización permite la producción de una lámina ungueal dura, córnea y transparente, compuesta de células completamente queratinizadas, aplanadas y enormemente adherentes entre sí, que han perdido completamente sus núcleos. La matriz se divide en tres capas: dorsal o superficial, intermedia o matriz real, y ventral o profunda, contribuyendo, cada una, a la formación de la lámina ungueal, en mayor o menor grado (Fig. 11).

Figura 11. Formación de la uña en la matriz ungueal.
Cualquier daño severo en la matriz, incluyendo procedimientos quirúrgicos, puede acarrear trastornos en el crecimiento ungueal, o incluso distrofias permanentes. Es, por ello, importante conocer su extensión anatómica. Está situada bajo el pliegue ungueal proximal. Se inicia en la lúnula y llega hasta una línea virtual ubicada en la mitad de la distancia entre cutícula y el pliegue de la articulación interfalángica distal. Y es la principal responsable del crecimiento ungueal, aportando el 81% de las células de la lámina ungueal.
La matriz contiene melanocitos que habitualmente no producen melanina. No obstante, pueden, en un momento dado, ser activados y sintetizar pigmento que transferirían a los queratinocitos de alrededor. La migración distal de queratinocitos conteniendo melanina, daría lugar a una lámina ungueal pigmentada.
La porción distal de la matriz ungueal es visible a través de la lámina ungueal transparente, como una media luna convexa distalmente: es la lúnula. Se encuentra en estrecho contacto con el pliegue ungueal proximal y es más visible en los primeros dedos de pies y manos. Su color más blanquecino, no es debido a ninguna diferencia de color con el lecho ungueal subyacente, sino porque la lúnula está menos adherida al citado lecho, la superficie de la uña es más suave a ese nivel y posee menos capilares en la dermis subyacente.
Toda esta estructura ungueal (Fig. 12) descansa sobre un área especializada de la falange distal, que condiciona la forma de la uña: las no infrecuentes exóstosis a ese nivel, favorecen el desarrollo de uña en pinza, o las falanges estrechas/cortas se asocian a uña en raqueta.
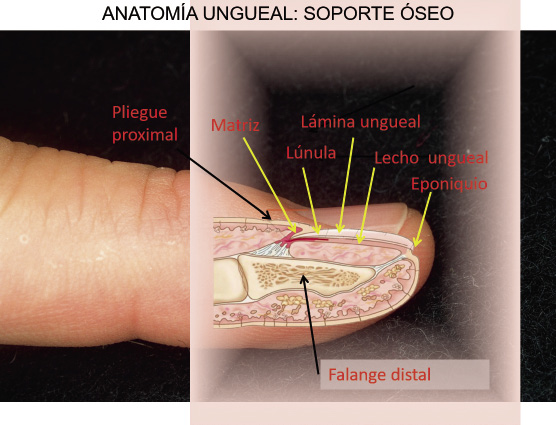
Figura 12. Anatomía ungueal: soporte óseo.
La vascularización de la región ungueal se realiza de forma abundante a través de unas arteriolas derivadas de las arterias digitales. Se forman dos arcos, uno superficial a la altura del repliegue ungueal (arco proximal dorsal) y otro profundo, por debajo del lecho ungueal (arco dorsal distal). Desde estas estructuras, se dividen una gran cantidad de arteriolas y capilares en forma de asas y anastomosis glómicas, que permiten la importante irrigación de la zona.
Un sistema venoso superficial compuesto por venas digitales palmares y dorsales, y otro profundo, son los responsables del drenaje venoso. La funcionalidad de los capilares periungueales, su morfología y la arquitectura de los mismos, puede evaluarse mediante dermatoscopia. Supone una aportación diagnóstica importante en procesos patológicos que afecten a la microcirculación (esclerosis sistémica, lupus, dermatomiositis, fenómeno de Raynoud…).
La inervación viene facilitada por el nervio digital y sus tres fascículos que se extienden por lecho, matriz ungueal y pulpejos.
Una estructura peculiar localizada a nivel del lecho ungueal (de 93 a 501 por cm2) son los denominados cuerpos glómicos: anastomosis arteriovenosas y terminaciones nerviosas que configuran una estructura neurovascular implicada en la termorregulación y regulación del flujo capilar. Importante papel en preservar y mantener la circulación en situaciones de bajas temperaturas.
Fisiología del aparato ungueal
El aparato ungueal está implicado en funciones de diversa índole.
La uña es una estructura cuyo crecimiento puede ser medido fácilmente: el crecimiento diario de las uñas de la mano está entre 100 y 120 µm. Al mes, crecen entre 2-4 mm. El crecimiento en las uñas de los pies es la mitad aproximadamente. La tasa de crecimiento del dedo medio -de los tres dedos medios en general- es la más alta, mientras que el primer y quinto dedo muestran la menor tasa. La razón para ello es desconocida. El crecimiento ungueal es continuo a lo largo de toda la vida (a diferencia del folículo piloso que lo es de forma cíclica), comenzando en la semana 15 del desarrollo embrionario hasta la muerte, y diferente de una persona a otra. La matriz ungueal, actuando como un epitelio germinativo, se mantiene en continua actividad, lo que se traduce en proliferación y diferenciación de los queratinocitos. Varios factores fisiológicos y estados patológicos pueden influir en la tasa de crecimiento ungueal (Tabla III).

La queratinización de la lámina ungueal sigue un eje oblicuo (Fig. 11), ya que durante el proceso de maduración y diferenciación, las células se mueven hacia arriba y distalmente. Por esta razón, la porción proximal de la matriz ungueal produce la parte más superior de la lámina ungueal, mientras que la porción más distal de la citada matriz es responsable de la zona dorsal de la lámina ungueal. El pliegue ungueal proximal es el encargado de “reconducir” este crecimiento lineal, moldeando la uña a medida que se genera en la matriz.
La curvatura tan especial que esto produce, tanto en su eje longitudinal como en el transversal, condiciona unas peculiares propiedades físicas a la lámina ungueal. Desde el punto de vista práctico, es interesante el papel de la permeabilidad, ya que condiciona la penetración de los diversos medicamentos: tópicos (a través de la superficie dorsal) como sistémicos (a través de la superficie ventral). La permeabilidad también dependerá de factores ajenos a la lámina, como es el tamaño de la molécula que la intenta atravesar. Además de la composición innata de la uña, pueden depositarse en ellas productos exógenos (medicamentos, tóxicos, drogas), capaces de producir efectos deletéreos en el desarrollo ungueal normal. Lo que conocemos como configuración ungueal normal, es el resultado de la interactuación de una serie de factores: genéticos, nutricionales, vasculares, neurogénicos y mecánicos.
Resto de anejos
Las características particulares de cada uno de los restantes anejos, quedan recogidas en los esquemas correspondientes: pelo (Fig. 13), glándulas sebáceas (Fig. 14) y glándulas sudoríparas ecrinas y apocrinas (Fig. 15).

Figura 13. Pelo – vello. Estructuras queratinizadas que asientan en toda la piel a excepción de palmas, plantas, labios, pezones, partes de genitales externos y extremos distales de los dedos. Estructura: en el tallo (A) se observan tres capas concéntricas: médula del pelo, corteza y cutícula. El músculo erector del pelo (B) se origina de la dermis adyacente al folículo piloso y tiene una dirección oblicua. En la parte inferior de la raíz folicular (C), se localiza el bulbo piloso (D), donde se encuentran las células epidérmicas que dan origen al pelo y rodean a la papila dérmica (E), que contiene capilares y nutre a las células epidérmicas. Entre estas células epidérmicas se encuentran melanocitos que dan color al cabello. En la raíz, se observan células epidérmicas que forman la vaina radicular (F) interna y otras más periféricas que forman la vaina radicular externa.
Fases del pelo: fase de crecimiento o “anágeno” en la que se encuentra: 80% cabello y dura entre 2-5 años; fase de transición o “catágeno”, fase más corta, de unos 14 días, en la que se produce la detención de la mitosis; fase de reposo o “telógeno” en la que está: 20% cabello, tiene una duración de 3 meses y finaliza en la caída del cabello.

Figura 14. Glándulas sebáceas. Glándulas holocrinas que producen lípidos que ayudan a mantener el manto hidrolipídico de la piel.
Localización: localizadas en toda la piel, excepto en palmas y plantas. Más abundantes en cara y cuero cabelludo.
Morfología: su conducto excretor desemboca en el folículo piloso, excepto en determinadas áreas en que no se relacionan con el folículo y drenan directamente a la superficie cutánea: areola, pezón (tubérculos de Montgomery), labios (manchas de Fordyce), labios menores y glande (glándulas de Tyson), y párpados (glándulas de Meibomio).
Actividad: la aumentan en la pubertad. Su número y grado de actividad son muy variables entre las diferentes personas. Desarrollo: se detecta su presencia a las 16-18 semanas de gestación y se completa en el tercer trimestre. Contribuyen en esta fase intraútero a la formación del vérnix, un manto elaborado de sebo y células descamadas, presente ya en el tercer trimestre gestacional. En los nacidos muy prematuros, puede ser inexistente.
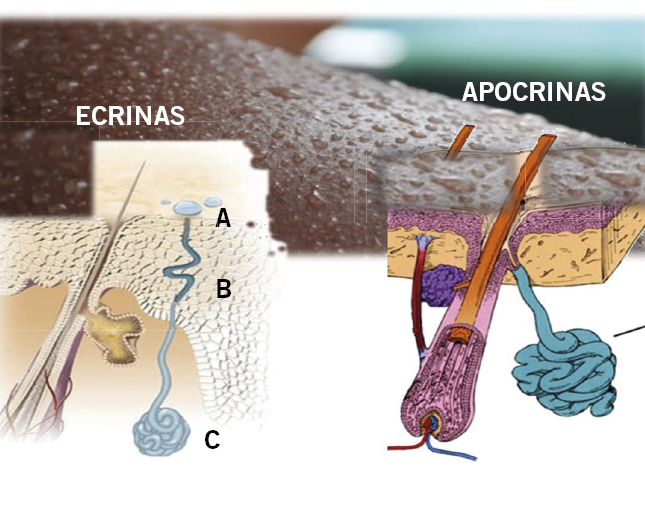
Figura 15. Glándulas sudoríparas. Las ecrinas son las más importantes. Las apocrinas tienen escasa representación en el hombre. Son glándulas tubulares, ubicadas en dermis, que forman un glomérulo u ovillo en su extremo. Los niños antes de la pubertad tienen un olor diferente a los adultos, ya que no producen sudor apocrino. De todo el aparato secretor ecrino, las glándulas palmoplantares son las primeras en desarrollarse. A las 22 semanas, ya se abren a la superficie palmoplantar y, entre las 24-26, se completa la maduración. La sudoración inducida por el estímulo térmico no está presente al nacimiento, pero suele ya estarlo a las dos semanas de vida.
Ecrinas. Producen el sudor y con ello regulan la temperatura. Se encuentran localizadas en todo el cuerpo, habiendo una mayor cantidad en palmas y plantas. Estructura: porción secretora o glomérulo (C). Porción excretora que drena directamente a la superficie cutánea: intraepidérmica o acrosiringio (A) y dérmica o ducto dérmico (B). Responden a estímulos tales como: calor, ciertos alimentos, situaciones de estrés y estímulos simpáticos, y para simpáticos.
Apocrinas. Son glándulas profundas localizadas a nivel de dermis dprofunda o tejido graso subcutáneo y que drenan directamente al folículo piloso. Localización: mayoritariamente en: axila, área anogenital, periumbilical, párpados (glándulas de Moll), areola y pezón. Función odorífera. El olor de su producción se debe a la degradación de su secreción por la flora bacteriana. El mecanismo de secreción es apocrina o por decapitación. Responden a estímulos adrenérgicos y colinérgicos.
Bibliografía
1. Kim JY, Dao H. Physiology, Integument. In: StatPearls (Internet). Treasure Island (FL): StatPearls Publishing; 2020.
2. Hoeger PH. Physiology of Neonatal Skin. En: Harper’s Textbook of Pediatric Dermatology, Fourth Edition. Chapter 444. Editorial Wiley-Blackwell. 2019.
3. de-Souza IMF, Vitral GLN, Reis ZSN. Skin thickness dimensions in histological section measurement during late-fetal and neonatal developmental period: A systematic review. Skin Res Technol. 2019; 25: 793-800.
4. Liu Q, Zhang Y, Danby SG, Cork MJ, Stamatas GN. Infant Skin Barrier, Structure, and Enzymatic Activity Differ from Those of Adult in an East Asian Cohort. Biomed Res Int. 2018; 18: 1302465.
5. Pastushenko I, Prieto-Torres L, Gilaberte YC. Blanpain C. Células madre de la piel: en la frontera entre el laboratorio y la clínica. Parte I: células madre epidérmicas. Actas Dermosifiliogr. 2015; 106: 725-32.
6. Jurica SA, ?oli? A, Gveri?-Ahmetaševi? S, Lon?arevi? D, Filipovi?-Gr?i? B, Stipanovi? J, et al. Skin of the very premature newborn – physiology and care. Paediatr Croat. 2016; 60: 21-6.
7. Cracowski JL, Roustit M. Human Skin Microcirculation. Compr Physiol. 2020; 10: 1105-54.
8. Taïeb A. Skin barrier in the neonate. Pediatr Dermatol. 2018; 35: s5-s9.
9. Walters RM, Khanna P, Chu M, Mack MC. Developmental Changes in Skin Barrier and Structure during the First 5 Years of Life. Skin Pharmacol Physiol. 2016; 29: 111-8.
10. Kabashima K, Honda T, Ginhoux F, Egawa G. The immunological anatomy of the skin. Nat Rev Immunol. 2019; 19: 19-30.
11. Nguyen AV, Soulika AM. The Dynamics of the Skin’s Immune System. Int J Mol Sci. 2019; 20: 1811.
12. Pereira Garzón CM, Morales Cardona CA. Biología de la uña y su aplicación clínica. Piel. 2016. 31: 685-95.
13. Martín B. Histopatología de la uña. Actas Dermosifiliogr. 2013; 104: 564-78.
14. De Berker D. Nail anatomy. Clin Dermatol. 2013; 31: 509-15.

 Atopic dermatitis and seborrheic dermatitis
Atopic dermatitis and seborrheic dermatitis